Organización molecular de la membrana.
Para llevar a cabo las reacciones químicas necesarias en el mantenimiento de la vida, la célula necesita mantener un medio interno apropiado. Esto es posible porque las células se encuentran separadas del mundo exterior por una membrana limitante, la membrana plasmática. Además, la presencia de membranas internas en las células eucariotas proporciona compartimientos adicionales que limitan ambientes únicos en los que se llevan al cabo funciones altamente específicas, necesarias para la supervivencia celular.
La membrana plasmática se encarga de:
- Aislar selectivamente el contenido de la célula del ambiente externo.
- Regular el intercambio de sustancias entre el interior y exterior celular (lo que entra y sale de la célula).
- Comunicación intercelular.
Nota: La mayoría de las células tienen membranas internas además de la membrana plasmática, forman y delimitan compartimentos donde se llevan a a cabo las actividades bioquímicas de la célula. Las restantes membranas también constituyen barreras selectivas para el pasaje de sustancias.
Estructura de la Membrana:
Hacia 1935 Danielli y Davson sintetizaron los conocimientos proponiendo que la membrana plasmática estaba formaba por una "bicapa lipídica" con proteínas adheridas a ambas caras de la misma.
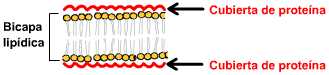
La integración de los datos químicos, físico-químicos y las diversas técnicas de microscopía llevó al actual modelo de "mosaico fluido" (Singer S.J., and Nicolson, G.L. (1972 Science, 175:120). Según este modelo del mosaico fluido, que ha tenido gran aceptación, las membranas constan de una bicapa lipídica (una doble capa de lípidos) en la cual están inmersas diversas proteínas.
Video Fluid mosiac model: http://www.youtube.com/watch?v=ULR79TiUj80
La bicapa lipídica ha sido establecida como la base universal de la estructura de la membrana celular y en ella sus componentes pueden realizar movimientos de traslación. Esta fluidez implica que los componentes en su mayoría solo están unidos por uniones no covalentes.
Los lípidos constituyen aproximadamente el 50% de la masa de la mayoría de las membranas plasmáticas de las células animales. La molécula primaria de la membrana celular es el fosfolípido, el cual posee una "cabeza" polar (hidrofílica) y dos "colas" no polares (hidrofóbicas), por tanto son andipáticos.
El colesterol es otro componente importante de la membrana. Se encuentra embebido en el área hidrofóbica de la misma, su presencia contribuye a la estabilidad de la membrana.
Proteínas de la membrana
Las proteínas de la membrana pueden considerarse, de acuerdo a como se encuentran en la membrana, comprendidas en una de estas dos categorías:
La superficie externa de la membrana tiende a ser rica en glicolípidos que tienen su colas hidrofóbicas embebidas en la región hidrofóbica de la membrana y sus cabezas hacia el exterior de la célula. Ellos, junto a con los hidratos de carbono pegados a las proteínas (glicoproteínas), intervienen en el reconocimiento de lo propio ("self") de un organismo. Los antígenos de diferenciación, mas conocidos como antígenos CD (por Cluster of Differentiation, grupo de diferenciación) no son otra cosa que glicoproteínas que se expresan sobre la superficie de las membranas. |

Esquema que representa a la membrana celular y sus componentes.
La membrana plasmática no es una estructura estática, sus componentes tienen posibilidades de movimiento, lo que le proporciona una cierta fluidez.
Los movimientos que pueden realizar los lípidos son:
1. De rotación: es como si girara la molécula en torno a su eje. Es muy frecuente y el responsable en parte de los otros movimientos.
2. De difusión lateral: las moléculas se difunden de manera lateral dentro de la misma capa. Es el movimiento más frecuente.
3. Flip-flop: es el movimiento de la molécula lipídica de una monocapa a la otra gracias a unas enzimas llamadas flipasas. Es el movimiento menos frecuente, por ser energéticamente más desfavorable
4. De flexión: son los movimientos producidos por las colas hidrófobas de los fosfolípidos.
La fluidez es una de las características más importantes de las membranas depende de factores como:
1. La temperatura, la fluidez aumenta al aumentar la temperatura.
2. La naturaleza de los lípidos, la presencia de lípidos insaturados y de cadena corta favorecen el aumento de fluidez; la presencia de colesterol endurece las membranas, reduciendo su fluidez y permeabilidad.
Transporte de moléculas:
Lo primero que tiene que hacer un microorganismo a la hora de su nutrición es captar los nutrientes que necesite desde el medio exterior. Debido a que la bicapapa lipídica actúa como barrera que impide el paso de la mayor parte de las sustancias, esto significa que deben existir mecanismos específicos para lograr la entrada de los nutrientes. Además, teniendo en cuenta que las bacterias suelen vivir en medios diluidos, deben realizar un “trabajo” para trasladar muchos de esos nutrientes en contra del gradiente de concentración.
Tradicionalmente se han considerando tres métodos principales de transporte de sustancias a través de la membrana:
- Transporte pasivo inespecífico (= difusión simple); consiste en la difusión de ciertas sustancias para las que la membrana es impermeable, debido a la diferencia de concentración (DC) a ambos lados de dicha membrana (la sustancia tiene mayor concentración fuera que dentro de la célula). Las membranas citoplásmicas son impermeables en sí mismas a la mayor parte de las moléculas. La difusión simple se produce por el paso de estas sustancias a través de poros inespecíficos de la membrana citoplásmica.
- Transporte pasivo específico (= difusión facilitada); es un proceso que permite el paso de compuestos por difusión a través de transportadores estereoespecíficos. El transportador suele ser una proteína integral de membrana (permeasa o facilitador), cuya conformación determina un canal interior, y por el cual un determinado sustrato puede alcanzar el interior, sin gasto de energía.
Video de Transporte Pasivo: http://www.youtube.com/watch?v=s0p1ztrbXPY&NR=1
- Transporte activo; consiste en el transporte de sustancias en contra de un gradiente de concentración, lo que requiere un gasto energético. En la mayor parte de los casos este transporte activo (que supone un trabajo osmótico) se realiza a expensas de un gradiente de H+ (potencial electroquímico de protones) previamente creado a ambos lados de la membrana, por procesos de respiración y fotosíntesis; y por hidrólisis de ATP.
Video de Transporte Activo: http://www.youtube.com/watch?v=STzOiRqzzL4&NR=1
| Esquema de comparación entre transporte activo y pasivo |
Video de membrana plasmática: http://www.youtube.com/watch?v=moPJkCbKjBs&feature=related
La Matriz Extracelular
Las interacciones celulares resultan fundamentales para su integración en tejidos y su relación con células similares o diferentes.
Las células animales secretan alrededor de ellas un complejo retículo conformado por proteínas e hidratos de carbono que les crean un ambiente especial: la matriz extracelular. Sus principales componentes son:
- El colágeno, fibras proteicas que confieren resistencia y fortaleza a la matriz.
- Los proteoglucanos, glicoproteínas que poseen una proporción de polisacáridos mayor que lo usual. y confieren el alto grado de viscosidad característico de la matriz.
- Las fibronectinas, proteínas multiadhesivas, tienen afinidad tanto para el colágeno como para las integrinas de las células. Su función principal es la fijación de células a matrices que contienen colágeno.
Nota: La matriz extracelular de las células animales puede equipararse a la pared celular de las células vegetales, cuya composición química es muy diferente y se describe mas adelante.
Las funciones principales de la matriz extracelular son:
- Rellenar los intersticios o espacios entre las células.
- Conferir resistencia mecánica (a la compresión, estiramiento, etc.) a los tejidos.
- Constituir el medio homeostático, nutritivo y metabólico para las células.
- Proveer fijación para el anclaje celular.
- Constituir el medio táctico para el tránsito celular, principalmente en la organogénesis.
- Comunicación celular: Ser el medio por el cual se transportan diferentes señales entre las células.
- Ser un reservorio de diferentes hormonas
Adhesión intercelular
Un grupo de proteínas denominadas Moléculas de Adhesión Celular (MAC o CAM por sus siglas en inglés) es el responsable de las interacciones entre células. Estas proteínas corresponden a proteínas integrales de membrana , entre las mas importantes tenemos:
- Cadherinas: son responsables de las interacciones entre células similares (interacciones homotípicas) y requieren de Ca ++ para dicha interacción, y entre otras características , se encuentra las de relacionarse (por su porción citosólica) al citoesqueleto Un ejemplos de uniones que contiene cadherina lo constituyen los desmosomas y las uniones adhesivas celulares y que confieren rigidez y fortaleza al conjunto de células que se unen para formar tejidos.
- Selectinas: son responsables de las interacciones entre células diferentes (interacciones heterótípicas), se fijan a los hidratos de carbono de otras moléculas de adhesión celular. Esta fijación es Ca++ dependiente y se realiza por medio de una lectina que se encuentra en el extremo de la molécula.


Esquema simplificado de uniones que se establecen entre células.
- Los desmosomas (como se observa en la figura superior), constan de una placa adosada a la cara citosólica de las respectivas membranas citoplasmáticas de las células que unen y, siendo las cadherinas los elementos que unen a las mismas. La placa (formada por proteínas denominadas placoglobinas) se unen a filamentos intermedios del citoesqueleto (queratina). Las cadherinas (en este caso proteínas de trasmembrana denominadas desmogleína y desmocolina) se fijan a la placa y se proyectan al espacio intercelular entrelazándose a las de la otra célula.
- Uniones adherentes: se las encuentra generalmente en el tejido epitelial conformando una banda continua de moléculas de cadherina que en su porción citosólica se unen a un "cinturón" de proteínas adaptadoras que discurre en la cara citosólica de la membrana celular y relaciona a las cadherinas con el citoesqueleto (principalmente actina).
- Unión estrecha u oclusiva: Se las encuentra separando los líquidos extracelulares que bañan las regiones apicales y basales de las células (con el objeto de que cumplan sus respectivas funciones) y forman barreras que tornan impermeables determinadas cavidades (como la luz del intestino). En este tipo de relación entre células, hileras de proteínas integrales de membrana (como la ocludina y la claudina) forman, con la porción que se proyecta al espacio intercelular, uniones extremadamente fuertes con las similares de la célula adyacente y prácticamente fusionan ambas células estableciendo una unión impermeable. La porción citosólica de estas células se relaciona al citoesqueleto.
Comunicación intercelular

Uniones comunicantes (gap junctions)
Un tipo particular de unión entre células animales lo constituye la unión comunicante (gap junction), en este caso las membranas de ambas poseen proteínas que conforman semicanales de transmembrana, que las interconectan y permiten el paso de moléculas entre ambas.
NOTA: En las células vegetales las uniones intercelulares se extienden a través de las paredes celulares de células adyacentes y se denominan plasmodesmos. Al igual que las uniones comunicantes, conectan a ambas células permitiendo el paso de moléculas, pero en este caso la membrana celular conforma una lámina continua que "tapiza" el plasmodesmo y, por otra parte una extensión del retículo plasmático (el desmotúbulo) lo atraviesa y se conecta al citosol de la célula adyacente.
Biosíntesis de colágena:
El colágeno es una molécula proteica que forma fibras; estas se encuentran en todos los animales pluricelulares y son secretadas por las células del tejido conjuntivo como los fibroblastos, así como por otros tipos celulares. Es el componente más abundante de la piel y de los huesos, cubriendo un 25% de la masa total de proteínas en los mamíferos. Cada una de las cadenas polipeptídicas es sintetizada por los ribosomas unidos a la membrana del retículo endoplásmico y luego son traslocadas al lumen del mismo en forma de grandes precursores (procadenas α), presentando aminoácidos adicionales en los extremos amino y carboxilo terminales. En el retículo endoplásmico los residuos de prolina y lisina son hidroxilados para luego algunos ser glucosilados en el aparato de Golgi; parece ser que estas hidroxilaciones son útiles para la formación de puentes de hidrógeno intercatenarios que ayudan a la estabilidad de la superhélice. Tras su secreción, los propéptidos de las moléculas de procolágeno son degradados mediante proteasas convirtiéndolas en moléculas de tropocolágeno asociándose en el espacio extracelular formando las fibrillas de colágeno. La formación de fibrillas está dirigida, en parte, por la tendencia de las moléculas de procolágeno a autoensamblarse mediante enlaces covalentes entre los residuos de lisina, formando un empaquetamiento escalonado y periódico de las moléculas de colágeno individuales en la fibrilla.

Esquema de la Síntesis de Colágeno-Formación de fibras de colágeno.
Adhesión entre las células y la matriz
En los grupos organizado de células la matriz, entre otras funciones, cumple la de organizar las células en tejidos amén de coordinarlas proporcionando el medio para que se propaguen señales que pueden indicar a las células que crezcan y proliferen.
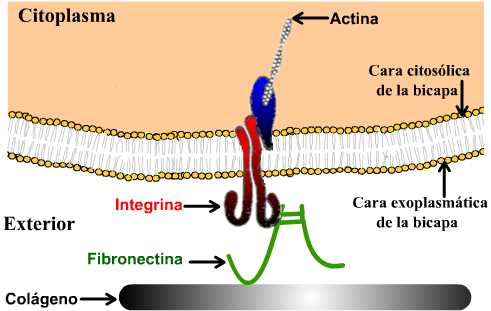
Esquema simplificado de la matriz celular, que muestra una de las relaciones entre componentes de la matriz y componentes del citoesqueleto.
La adhesión entre las células y la matriz esta mediada esencialmente por las integrinas: son las principales clases de MAC que interaccionan entre la célula y la matriz (aunque las selectinas y proteoglucanos también intervienen en la fijación). Las integrinas están compuestas por dos subunidades diferentes (heterodímeros) que toman el nombre de alfa (con 17 tipos diferentes) y beta (con ocho tipos diferentes), lo cual permite un gran número de combinaciones. Las células generalmente presentan en su superficie varios tipos de integrinas.
La porción extracelular de la integrina se fija a las proteínas de la matriz y la citosólica se relaciona con proteínas adaptadoras que a su vez interactúan con el citoesqueleto. Algunas integrinas pueden además de mediar entre la célula y la matriz, intervenir en interacciones intercelulares.
La Matriz Extracelular
- El colágeno, fibras proteicas que confieren resistencia y fortaleza a la matriz.
- Los proteoglucanos, glicoproteínas que poseen una proporción de polisacáridos mayor que lo usual. y confieren el alto grado de viscosidad característico de la matriz.
- Las fibronectinas, proteínas multiadhesivas, tienen afinidad tanto para el colágeno como para las integrinas de las células. Su función principal es la fijación de células a matrices que contienen colágeno.
Adhesión intercelular
Un grupo de proteínas denominadas Moléculas de Adhesión Celular (MAC o CAM por sus siglas en inglés) es el responsable de las interacciones entre células. Estas proteínas corresponden a proteínas integrales de membrana , entre las mas importantes tenemos:
- Cadherinas: son responsables de las interacciones entre células similares (interacciones homotípicas) y requieren de Ca ++ para dicha interacción, y entre otras características , se encuentra las de relacionarse (por su porción citosólica) al citoesqueleto Un ejemplos de uniones que contiene cadherina lo constituyen los desmosomas y las uniones adhesivas celulares y que confieren rigidez y fortaleza al conjunto de células que se unen para formar tejidos.
- Selectinas: son responsables de las interacciones entre células diferentes (interacciones heterótípicas), se fijan a los hidratos de carbono de otras moléculas de adhesión celular. Esta fijación es Ca++ dependiente y se realiza por medio de una lectina que se encuentra en el extremo de la molécula.
 |
Esquema simplificado de uniones que se establecen entre células. |
| |
| |
|
Comunicación intercelular
|
| Uniones comunicantes (gap junctions) |
Un tipo particular de unión entre células animales lo constituye la unión comunicante (gap junction), en este caso las membranas de ambas poseen proteínas que conforman semicanales de transmembrana, que las interconectan y permiten el paso de moléculas entre ambas.
NOTA: En las células vegetales las uniones intercelulares se extienden a través de las paredes celulares de células adyacentes y se denominan plasmodesmos. Al igual que las uniones comunicantes, conectan a ambas células permitiendo el paso de moléculas, pero en este caso la membrana celular conforma una lámina continua que "tapiza" el plasmodesmo y, por otra parte una extensión del retículo plasmático (el desmotúbulo) lo atraviesa y se conecta al citosol de la célula adyacente.
Biosíntesis de colágena:
El colágeno es una molécula proteica que forma fibras; estas se encuentran en todos los animales pluricelulares y son secretadas por las células del tejido conjuntivo como los fibroblastos, así como por otros tipos celulares. Es el componente más abundante de la piel y de los huesos, cubriendo un 25% de la masa total de proteínas en los mamíferos.
Cada una de las cadenas polipeptídicas es sintetizada por los ribosomas unidos a la membrana del retículo endoplásmico y luego son traslocadas al lumen del mismo en forma de grandes precursores (procadenas α), presentando aminoácidos adicionales en los extremos amino y carboxilo terminales. En el retículo endoplásmico los residuos de prolina y lisina son hidroxilados para luego algunos ser glucosilados en el aparato de Golgi; parece ser que estas hidroxilaciones son útiles para la formación de puentes de hidrógeno intercatenarios que ayudan a la estabilidad de la superhélice.
Tras su secreción, los propéptidos de las moléculas de procolágeno son degradados mediante proteasas convirtiéndolas en moléculas de tropocolágeno asociándose en el espacio extracelular formando las fibrillas de colágeno.
La formación de fibrillas está dirigida, en parte, por la tendencia de las moléculas de procolágeno a autoensamblarse mediante enlaces covalentes entre los residuos de lisina, formando un empaquetamiento escalonado y periódico de las moléculas de colágeno individuales en la fibrilla.
 |
| Esquema de la Síntesis de Colágeno-Formación de fibras de colágeno. |
Adhesión entre las células y la matriz
En los grupos organizado de células la matriz, entre otras funciones, cumple la de organizar las células en tejidos amén de coordinarlas proporcionando el medio para que se propaguen señales que pueden indicar a las células que crezcan y proliferen.
| ||
La adhesión entre las células y la matriz esta mediada esencialmente por las integrinas: son las principales clases de MAC que interaccionan entre la célula y la matriz (aunque las selectinas y proteoglucanos también intervienen en la fijación). Las integrinas están compuestas por dos subunidades diferentes (heterodímeros) que toman el nombre de alfa (con 17 tipos diferentes) y beta (con ocho tipos diferentes), lo cual permite un gran número de combinaciones. Las células generalmente presentan en su superficie varios tipos de integrinas.
No hay comentarios:
Publicar un comentario